Торможение виды – 23. Виды торможения в цнс. Тормозные нейроны и тормозные синапсы. Торможение с участием и без участия специфических тормозных структур.
- 29.06.2020
Торможение (физиология) — Википедия
Материал из Википедии — свободной энциклопедии
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 февраля 2018; проверки требуют 8 правок. Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 февраля 2018; проверки требуют 8 правок.Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
И. П. Павлов называл иррадиацию торможения по коре больших полушарий головного мозга «проклятым вопросом физиологии».
Центральное торможение (Виды торможения)[править | править код]
Центральное торможение открыто в 1862 г. И. М. Сеченовым. В процессе опыта он удалил у лягушки головной мозг на уровне зрительных бугров и определял время сгибательного рефлекса. Затем на зрительные бугры помещался кристалл соли в результате чего наблюдалось увеличение продолжительности времени рефлекса. Это наблюдение позволило И. М. Сеченову высказать мнение о явлении торможения в ЦНС. Данный тип торможения называют
Ухтомский объяснил результаты с позиции доминанты. В зрительных буграх — доминанта возбуждения, которая подавляет действие спинного мозга.
Введенский объяснил результаты с позиции отрицательной индукции. Если в центральной нервной системе возникает возбуждение в определённом нервном центре, то вокруг очага возбуждения индуцируется торможение.
Современное объяснение: при раздражении зрительных бугров возбуждается каудальный отдел ретикулярной формации. Эти нейроны возбуждают тормозные клетки спинного мозга (клетки Реншоу), которые тормозят активность альфа-мотонейронов спинного мозга.
Первичное торможение[править | править код]
Первичное торможение возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы.
Виды первичного торможения
- Постсинаптическое — основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Примеры первичного торможения:
- Возвратное — нейрон воздействует на клетку, которая, в ответ тормозит этот же нейрон.
- Реципрокное — это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон.
- Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клетками сетчатки, что создаёт условия для более четкого видения предмета.
- Возвратное облегчение — нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками.
- Пресинаптическое
Вторичное торможение[править | править код]
Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
Виды вторичного торможения
- Пессимальное торможение — это вторичное торможение, которое развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под действием множественной импульсации.
- Торможение вслед за возбуждением возникает в обычных нейронах и также связано с процессом возбуждения. В конце акта возбуждения нейрона в нем может развиваться сильная следовая гиперполяризация. В то же время возбуждающий постсинаптический потенциал не может довести деполяризацию мембраны до критического уровня деполяризации, потенциалзависимые натриевые каналы не открываются и потенциал действия не возникает.
Открыто братьями Вебер в 1845 г. В качестве примера можно привести торможение деятельности сердца (снижение ЧСС) при раздражении блуждающего нерва.
Термины «условное» и «безусловное» торможение предложены И. П. Павловым.
Условное торможение[править | править код]
Условное, или внутреннее, торможение — форма торможения условного рефлекса, возникающее при неподкреплении условных раздражителей безусловными. Условное торможение является приобретённым свойством и вырабатывается в процессе онтогенеза. Условное торможение является центральным торможением и ослабевает с возрастом[1].
Безусловное торможение[править | править код]
Безусловное (внешнее) торможение — торможение условного рефлекса, возникающее под действием безусловных рефлексов (например, ориентировочного рефлекса). И. П. Павлов относил безусловное торможение к врождённым свойствам нервной системы, то есть безусловное торможение является формой центрального торможения[1].
- ↑ 1 2 главный редактор Гиляров М. С. Биология. Большой энциклопедический словарь. — 3. — Москва: Большая Российская Энциклопедия, 1999. — 864 с. — 100 000 экз. — ISBN 5852702528.
Торможение. Виды торможения. Биологическая значимость торможения (стр. 1 из 3)
Пермский гуманитарно-технологический институт
Гуманитарный факультет
КОНТРОЛЬНАЯ РАБОТА
По дисциплине «Физиология ВНД»
Тема «Торможение. Виды торможения. Биологическая значимость торможения»
Выполнил студент группы П-07-2з
Дмитрий Валерьевич
Проверила: Третьякова М.В.
Пермь, 2009
Содержание:
Введение
Торможение
Виды торможения
Значение торможения
Заключение
Список литературы
Введение
«Если бы животное не было… точно приспособлено к внешнему миру, то оно скоро или медленно перестало бы существовать… Оно так должно реагировать на внешний мир, чтобы всей ответной деятельностью его было обеспечено его существование». И.П. Павлов.
Приспособление животных и человека к изменяющимся условиям существования во внешней среде обеспечивается деятельностью нервной системы и реализуется через рефлекторную деятельность. Для обеспечения приспособления и адекватного поведения необходимы не только способность к выработке новых условных рефлексов и их длительное сохранение, но и возможность к устранению тех условно-рефлекторных реакций, необходимость в которых отсутствует. Исчезновение условных рефлексов обеспечивается процессами торможения.
Что же такое торможение? Какие существуют виды торможения? Для чего оно нужно? Попытаемся разобраться в этом на страницах контрольной работы.
Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
По И.П.Павлову, различают следующие формы коркового торможения: безусловное, условное и запредельное торможение.
Безусловное торможение
Этот вид торможения условных рефлексов возникает сразу в ответ на действие постороннего раздражителя, т.е. является врожденной, безусловной формой торможения. Безусловное торможение может быть внешним и запредельным. Внешнее торможение возникает под влиянием нового раздражителя, создающего доминантный очаг возбуждения, формирующего ориентировочный рефлекс. Биологическое значение внешнего торможения состоит в том, что, затормаживая текущую условно-рефлекторную деятельность, оно позволяет переключить организм на определение значимости и степени опасности нового воздействия.
Посторонний раздражитель, оказывающий тормозящее влияние на течение условных рефлексов, называется внешним тормозом. При многократном повторении постороннего раздражителя вызываемый ориентировочный рефлекс постепенно уменьшается, а затем исчезает и уже не вызывает торможения условных рефлексов. Такой внешний тормозящий раздражитель называется гаснущим тормозом. Если же посторонний раздражитель содержит биологически важную информацию, то он всякий раз вызывает торможение условных рефлексов. Такой постоянный раздражитель называется постоянным тормозом.
Биологическое значение внешнего торможения — обеспечение условий для более важного в данный момент ориентировочного рефлекса, вызванного экстренным раздражителем, и создание условий для его срочной оценки.
Условное торможение (внутреннее)
Оно возникает, если условный раздражитель перестает подкрепляться безусловным. Его называют внутренним, потому что оно формируется в структурных компонентах условного рефлекса. Условное торможение требует для выработки определенного времени. К этому виду торможения относятся: угасательное, дифференцировочное, условный тормоз и запаздывающее.
Угасательное торможение развивается в тех случаях, когда условный раздражитель перестает подкрепляться безусловным, при этом условная реакция постепенно исчезает. При первом предъявлении условного раздражителя без последующего подкрепления условная реакция проявляется как обычно. Последующие предъявления условного раздражителя без подкрепления начинают вызывать ориентировочную реакцию, которая затем угасает. Постепенно исчезает и условно-рефлекторная реакция.
Дифференцировочное торможение вырабатывается на раздражители, близкие по характеристике к условному раздражителю. Этот вид торможения лежит в основе различения раздражителей. С помощью этого торможения из сходных раздражителей выделяется тот, который будет подкрепляться безусловным раздражителем, т.е. биологически важный для организма. Например, на звук метронома с частотой 120 ударов в 1 мин у собаки выделяется слюна. Если теперь этому животному в качестве раздражителя предъявить звук метронома с частотой 60 ударов, но не подкреплять его, то в первых опытах этот раздражитель тоже вызывает отделение слюны. Но через некоторое время возникает диф-ференцировка этих двух раздражителей и на звук с частотой 60 ударов слюна выделяться перестает.
Условный тормоз — это разновидность дифференцировочно-го торможения. Возникает в том случае, если положительный условный раздражитель подкрепляется безусловным, а комбинация из условного и индифферентного раздражителей не подкрепляется. Например, условный раздражитель свет подкрепляется безусловным раздражителем, а комбинация свет и звонок не подкрепляется. Первоначально эта комбинация вызывает такой же условный ответ, но в дальнейшем она утрачивает свое сигнальное значение и на нее условная реакция возникать не будет, в то время как на изолированный условный раздражитель (свет) она сохраняется. Звонок же приобретает значение тормозного сигнала. Его подключение к любому другому условному раздражителю затормаживает проявление условного рефлекса.
Запаздывающее торможение характеризуется тем, что условная реакция на условный раздражитель возникает до действия безусловного раздражителя. При увеличении интервала между началом действия условного раздражителя и моментом подкрепления (до 2-3 мин) условная реакция все более и более запаздывает и начинает возникать непосредственно перед предъявлением подкрепления. Отставание условной реакции от начала действие условного раздражителя свидетельствует о выработке запаздывающего торможения, так как период торможения соответствует периоду запаздывания подкрепления.
Условное торможение дает возможность организму избавиться от большого количества лишних биологически нецелесо образных реакций. Внутреннее торможение (по П.К.Анохину) является результатом борьбы двух потоков возбуждений при их выходе на эффекторы. При угасательном торможении, например, пищевого слюноотделительного условного рефлекса — это поток возбуждений соответствующей пищевой реакции и поток возбуждений, характерный для биологически отрицательной реакции, возникающий при отсутствии подкрепления. Более сильное, доминирующее возбуждение реакции неудовлетворения тормозит менее сильное, пищевое возбуждение.
Запредельное торможение
Этот вид торможения отличается от внешнего и внутреннего по механизму возникновения и физиологическому значению. Оно возникает при чрезмерном увеличении силы или продолжительности действия условного раздражителя, вследствие того, что сила раздражителя превышает работоспособность корковых клеток. Это торможение имеет охранительное значение, так как препятствует истощению нервных клеток. По своему механизму оно напоминает явление «пессимума», которое было описано Н.Е.Введенским.
Запредельное торможение может вызываться действием не только очень сильного раздражителя, но и действием небольшого по силе, но длительного и однообразного по характеру раздражения. Это раздражение, постоянно действуя на одни и те же корковые элементы, приводит их к истощению, а следовательно, сопровождается возникновением охранительного торможения. Запредельное торможение легче развивается при снижении работоспособности, например, после тяжелого инфекционного заболевания, стресса, чаще развивается у пожилых людей.
Все виды условного торможения имеют огромное значение в жизни человека. Выдержка и самообладание, точное распознавание окружающих нас предметов и явлений, наконец, точность и четкость движений невозможны без торможения. Есть все основания полагать, что торможение основано не просто на угнетении условных рефлексов, а на выработке особых тормозных условных рефлексов. Центральным звеном таких рефлексов является тормозная нервная связь. Тормозной условный рефлекс нередко называют отрицательным в противоположность положительному условному рефлексу.
Торможение нежелательной реакции сопряжено с большой тратой энергии. Конкурирующие раздражения, а также другие причины, связанные с физическим состоянием организма, могут ослабить процесс торможения и привести к растормаживанию. При растормаживании проявляются действия, которые ранее устранялись процессами торможения.
Заключение
Функционирование условно-рефлекторного механизма базируется на двух основных нервных процессах: на процессе возбуждения и на процессе торможения. По мере становления и упрочения условного рефлекса возрастает роль тормозного процесса. Торможение является фактором, способствующим приспособлению организма к окружающим его условиям. Так же торможение ослабляет процессы возбуждения в нервной системе и обеспечивает стабильность её работы.
При отсутствии торможения, процессы возбуждения нарастали бы и накапливались, что неминуемо привело бы к разрушению нервной системы и гибели организма.
ПРАКТИЧЕСКАЯ ЧАСТЬ
МЫШЕЧНО-СУСТАВНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Исследуемый садится за кинематометр и закрывает глаза. Исследователь поочередно задает угол, который исследуемый должен в последующем воспроизвести на большой и малой шкале прибора. В
Торможение
Торможение — в физиологии — активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
И. П. Павлов называл иррадиацию торможения по коре больших полушарий головного мозга «проклятым вопросом физиологии».
Центральное торможение
Центральное
торможение открыто в 1862 г. И. М. Сеченовым.
В процессе опыта он удалил у лягушки
головной мозг на уровне зрительных
бугров и определял время сгибательного
рефлекса. Затем на зрительные бугры
помещался кристалл
Ухтомский объяснил результаты с позиции доминанты. В зрительных буграх — доминанта возбуждения, которая подавляет действие спинного мозга.
Введенский объяснил
результаты с позиции отрицательной
индукции. Если в центральной нервной
системе возникает возбуждение в
определенном нервном центре, то вокруг
очага возбуждения индуцируется
торможение. Современное объяснение:
при раздражении зрительных бугров
возбуждается каудальный отдел ретикулярной
формации. Эти нейроны возбуждают
тормозные клетки спинного мозга (
Первичное торможение
Первичное торможение возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы.
Виды первичного торможения
Постсинаптическое — основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Примеры первичного торможения:
Возвратное — нейрон воздействует на клетку, которая в ответ тормозит этот же нейрон.
Реципрокное — это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон.
Латеральное — тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клеткамисетчатки, что создает условия для более четкого видения предмета.
Возвратное облегчение — нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками.
Пресинаптическое — возникает в обычных нейронах, связано с процессом возбуждения.
Вторичное торможение
Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
Виды вторичного торможения
Пессимальное торможение — это вторичное торможение, которое развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под действием множественной импульсации.
Торможение вслед за возбуждением возникает в обычных нейронах и также связано с процессом возбуждения. В конце акта возбуждения нейрона в нем может развиваться сильная следовая гиперполяризация. В то же время возбуждающий постсинаптический потенциал не может довести деполяризацию мембраны до
Периферическое торможение
Открыто братьями Вебер в 1845 г. В качестве примера можно привести торможение деятельности сердца (снижение ЧСС) при раздражении блуждающего нерва.
Условное и безусловное торможение
Термины «условное» и «безусловное» торможение предложены И. П. Павловым.
Условное торможение
Условное,
или внутреннее, торможение — форма
торможения условного рефлекса, возникающее
при неподкреплении условных раздражителей
безусловными. Условное торможение
является приобретенным свойством и
вырабатывается в процессе онтогенеза.
Условное торможение является центральным
торможением и ослабевает с возрастом.
Безусловное торможение
Безусловное (внешнее) торможение — торможение условного рефлекса, возникающее под действием безусловных рефлексов (например, ориентировочного рефлекса). И. П. Павлов относил безусловное торможение к врожденным свойствам нервной системы, то есть безусловное торможение является формой центрального торможения.[1]
Координирующая функция локальных нейронных сетей помимо усиления может выражаться и в ослаблении слишком интенсивной активности нейронов за счет их торможения.
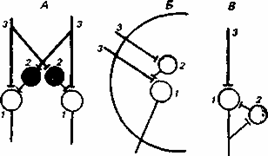
Рис.8.1 .Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга
1 — мотонейрон; 2 — тормозный интернейрон; 3 — афферентные терминали.
Торможение, как особый нервный процесс, характеризуется отсутствием способности к активному распространению по нервной клетке и может быть представлено двумя формами — первичным и вторичным торможением.
Первичное торможение обусловлено наличием специфических тормозных структур и развивается первично без предварительного возбуждения. Примером первичного торможения является так называемое реципрокное торможение мыщц-антагонистов, обнаруженное в спинальных рефлекторых дугах . Суть этого явления состоит в том, что если активируются проприорецепторы мышцы-сгибателя, то они через первичные афференты одновременно возбуждают мотонейрон данной мышцы-сгибателя и через коллатераль афферентного волокна тормозный вставочный нейрон. Возбуждение вставочного нейрона приводит к постсинаптическому торможению мотонейрона антагонистической мышцы-разгибателя, на теле которого аксон тормозного интернейрона формирует специализированные тормозные синапсы. Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
Другим примером первичного торможения является открытое Б. Реншоу возвратное торможение. Оно осуществляется в нейронной цепи, которая состоит из мотонейрона и вставочного тормозного нейрона — клетки Реншоу. Импульсы от возбужденного мотонейрона через отходящие от его аксона возвратные кол-латерали активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона. Это торможение реализуется за счет функции тормозных синапсов, которые клетка Реншоу образует на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов моторной клетки и подавить идущую к мышцам избыточную импульсацию.
В ряде случаев клетки Реншоу формируют тормозные синапсы не только на активирующих их мотонейронах, но и на соседних мотонейронах со сходными функциями. Осуществляемое через эту систему торможение окружающих клеток называется латеральным.
Торможение по принципу отрицательной обратной связи встречается не только на выходе, но и на входе моторных центров спинного мозга. Явление подобного рода описано в моносинаптических соединенях афферентных волокон со спинальными мотонейронами, торможение которых при данной ситуации не связано с изменениями в постсинаптической мембране. Последнее обстоятельство позволило определить данную форму торможения как пресинаптическое. Оно обусловлено наличием вставочных тормозных нейронов, к которым подходят коллатерали афферентных волокон . В свою очередь, вставочные нейроны формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению к мотонейронам. В случае избыточного притока сенсорной информации с периферии происходит активация тормозных интернейронов, которые через аксо-аксональные синапсы вызывают деполяризацию афферентных термина-леи и, таким образом, уменьшают количество выделяемого из них медиатора, а следовательно, и эффективность синаптической передачи. Электрофизиологическим показателем этого процесса является снижение амплитуды регистрируемых от мотонейрона ВПСП. Вместе с тем никаких признаков изменений ионной проницаемости или генерации ТПСП в мотонейронах не наблюдается.
Вопрос о механизмах пресинаптического торможения является довольно сложным. По-видимому, медиатором в тормозном аксо-аксональном синапсе является гамма-аминомасляная кислота, которая вызывает деполяризацию афферентных терминалей за счет увеличения проницаемости их мембраны для ионов С1-. Деполяризация снижает амплитуду потенциалов действия в афферентных волокнах и тем самым уменьшает квантовый выброс медиатора в синапсе. Другой возможной причиной деполяризации терминалей может быть повышение наружной концентрации ионов К+ при длительной активации афферентных входов. Следует отметить, что феномен пресинаптического торможения обнаружен не только в спинном мозгу, но и в других отделах ЦНС.
Исследуя координирующую роль торможения в локальных нейронных цепях, следует упомянуть еще об одной форме торможения — вторичном торможении, которое возникает без участия специализированных тормозных структур как следствие избыточной активации возбуждающих входов нейрона. В специальной литературе эту форму торможения определяют как торможение Введенского, который открыл его в 1886 г. при исследовании нервно-мышечного синапса.
Торможение Введенского играет предохранительную роль и возникает при чрезмерной активации центральных нейронов в полисинаптических рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны, превышающей критический уровень и вызывающей инактивацию Na-каналов, ответственных за генерацию потенциалов действия. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов импульсной активности нервных клеток.
ТОРМОЖЕНИЕ В ЦНС. ВИДЫ И ЗНАЧЕНИЕ.
Проявление и осуществление рефлекса возможно только при ограничении распространения возбуждения с одних нервных центров на другие. Это достигается взаимодействием возбуждения с другим нервным процессом, противоположным по эффекту процессом торможения.
Почти до середины XIX века физиологи изучали и знали только один нервный процесс — возбуждение.
Явления торможения в нервных центрах, т.е. в центральной нервной системе были впервые открыты в 1862 году И.М.Сеченовым («сеченовское торможение”). Это открытие сыграло в физиологии не меньшую роль, чем сама формулировка понятия рефлекса, так как торможение обязательно участвует во всех без исключения нервных актах. И.М.Сеченов обнаружил явление центрального торможения при раздражении промежуточного мозга теплокровных. В 1880 году немецкий физиолог Ф.Гольц установил торможение спинальных рефлексов. Н.Е. Введенский в результате серий опытов по парабиозу вскрыл интимную связь процессов возбуждения и торможения и доказал, что природа этих процессов едина.
Торможение —местный нервный процесс, приводящий к угнетению или предупреждению возбуждения. Торможение является активным нервным процессом, результатом которого служит ограничение или задержка возбуждения. Одна из характерных черт тормозного процесса- отсутствие способности к активному распространению по нервным структурам.
В настоящее время в центральной нервной системе выделяют два вида торможения:торможение центральное (первичное),являющееся результатом возбуждения (активации) специальных тормозных нейронов иторможение вторичное, которое осуществляется без участия специальных тормозных структур в тех самых нейронах в которых происходит возбуждение.
Центральное торможение(первичное) — нервный процесс, возникающий в ЦНС и приводящий к ослаблению или предотвращению возбуждения. Согласно современным представлениям центральное торможение связано с действием тормозных нейронов или синапсов, продуцирующих тормозные медиаторы (глицин, гаммааминомасляную кислоту), которые вызывают на постсинаптической мембране особый тип электрических изменений, названных тормозными постсинаптическими потенциалами (ТПСП) или деполяризацию пресинаптического нервного окончания, с которым контактирует другое нервное окончание аксона. Поэтому выделяют центральное (первичное) постсинаптическое торможение и центральное (первичное) пресинаптическое торможение.
Постсинаптическое торможение(лат. post позади, после чего-либо + греч. sinapsis соприкосновение, соединение) — нервный процесс, обусловленный действием на постсинаптическую мембрану специфических тормозных медиаторов (глицин, гаммааминомаслянная кислота), выделяемых специализированными пресинаптическими нервными окончаниями. Медиатор, выделяемый ими, изменяет свойства постсинаптической мембраны, что вызывает подавление способности клетки генерировать возбуждение. При этом происходит кратковременное повышение проницаемости постсинаптической мембраны к ионам К+ или CI, вызывающее снижение ее входного электрического сопротивления и генерацию тормозного постсинаптического потенциала (ТПСП). Возникновение ТПСП в ответ на афферентное раздражение обязательно связано с включением в тормозной процесс дополнительного звена — тормозного интернейрона, аксональные окончания которого выделяют тормозной медиатор. Специфика тормозных постсинаптических эффектов впервые была изучена на мотонейронах млекопитающих (Д. Экклс, 1951). В дальнейшем первичные ТПСП были зарегистрированы в промежуточных нейронах спинного и продолговатого мозга, в нейронах ретикулярной формации, коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
Известно, что при возбуждении центра сгибателей одной из конечностей центр ее разгибателей тормозится и наоборот. Д. Экклс выяснил механизм этого явления в следующем опыте. Он раздражал афферентный нерв, вызывающий возбуждение мотонейрона, иннервирующего мышцу — разгибатель.
Нервные импульсы, дойдя до афферентного нейрона в спинномозговом ганглии, направляются по его аксону в спинном мозге по двум путям: к мотонейрону, иннервирующему мышцу — разгибатель, возбуждая ее и по коллатерам к промежуточному тормозному нейрону, аксон которого контактирует с мотонейроном иннервирующим мышцу — сгибатель, вызывая таким образом торможение антагонистической мышцы. Этот вид торможения был обнаружении в промежуточных нейронах всех уровней центральной нервной системы при взаимодействии антагонистических центров. Он был назван поступательным постсинаптическим торможением. Этот вид торможения координирует, распределяет процессы возбуждения и торможения между нервными центрами.
Возвратное (антидромное) постсинаптическое торможение(греч. antidromeo бежать в противоположном направлении) — процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по принципу отрицательной обратной связи. Он заключается в том, что коллатерали аксонов нервной клетки устанавливают синаптические контакты со специальными вставочными нейронами (клетки Реншоу), роль которых заключается в воздействии на нейроны, конвергирующие на клетке, посылающей эти аксонные коллатерали (рис. 87). По такому принципу осуществляетсяторможение мотонейронов.
Возникновение импульса в мотонейроне млекопитающих не только активирует мышечные волокна, но через коллатерали аксона активирует тормозные клетки Реншоу. Последние устанавливают синаптические связи с мотонейронами. Поэтому усиление импульсации мотонейрона ведет к большей активации клеток Реншоу, вызывающей усиление торможения мотонейронов и уменьшение частоты их импульсации. Термин «антидромное” употребляется потому, что тормозной эффект легко вызывается антидромными импульсами, рефлекторно возникающими в мотонейронах.
Чем сильнее возбужден мотонейрон, чем больше сильные импульсы идут к скелетным мышцам по его аксону, тем интенсивнее возбуждается клетка Реншоу, которая подавляет активность мотонейрона. Следовательно, в нервной системе существует механизм, оберегающий нейроны от чрезмерного возбуждения. Характерная особенность постсинаптического торможения заключается в том, что оно подавляется стрихнином и столбнячным токсином (на процессы возбуждения эти фармакологические вещества не действуют).
В результате подавления постсинаптического торможения нарушается регуляция возбуждения в цнс, возбуждение разливается («диффундирует”) по всей цнс, вызывая перевозбуждение мотонейронов и судорожные сокращения групп мышц (судороги).
Торможение ретикулярное(лат. reticularis — сетчатый) — нервный процесс развивающийся в спинальных нейронах под влиянием нисходящей импульсации из ретикулярной формации (гигантское ретикулярное ядро продолговатого мозга). Эффекты, создаваемые ретикулярными влияниями, по функциональному действию сходны с возвратным торможением, развивающимся на мотонейронах. Влияние ретикулярной формации вызывают стойкие ТПСП, охватывающие все мотонейроны независимо от их функциональной принадлежности. В этом случае, так же как и при возвратном торможении мотонейронов происходит ограничение их активности. Между таким нисходящим контролем со стороны ретикулярной формации и системочй возвратного торможения через клетки Реншоу существует определенное взаимодействие, и клетки Реншоу находятся под постоянным тормозящем контролем со стороны двух структур. Тормозящее влияние со стороны ретикулярной формации являются дополнительным фактором в регуляции уровня активности мотонейронов.
Первичное торможение может вызываться механизмами иной природы, не связанными с изменениями свойств постсинаптической мембраны. Торможение в этом случае возникает на пресинаптической мембране (синаптическое и пресинаптическое торможение).
Синаптическое торможение(греч. sunapsis соприкосновение, соединение) — нервный процесс, основанный на взаимодействии медиатора, секретируемого и выделяемого пресинаптическими нервными окончаниями, со специфическими молекулами постсинаптической мембраны. Возбуждающий или тормозной характер действия медиатора зависит от природы каналов, которые открываются в постсинаптической мембране. Прямое доказательство наличия в цнс специфических тормозящих синапсов было впервые получено Д. Ллойдом (1941).
Данные относительно электрофизиологических проявлений синаптического торможения: наличие синаптической задержки, отсутствие электрического поля в области синаптических окончаний дали основание считать его следствием химического действия особого тормозящего медиатора, выделяемого синаптическими окончаниями. Д. Ллойд показал, что если клетка находится в состоянии деполяризации, то тормозной медиатор вызывает гиперполяризацию, в то время как на фоне гиперполяризации постсинаптической мембраны он вызывает ее деполяризацию.
Пресинаптическое торможение (лат. praе -впереди чего-либо + греч. sunapsis соприкосновение, соединение) — частный случай синаптических тормозных процессов, проявляющихся в подавлении активности нейрона в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким-либо изменениям. Пресинаптическое торможение осуществляется посредством специальных тормозных интернейронов. Его структурной основой являются аксо-аксональные синапсы, образованные терминалиями аксонов тормозных интернейронов и аксональными окончаниями возбуждающих нейронов.
При этом окончание аксона тормозного нейрона является пресимпатическим по отношению к терминали возбуждающего нейрона, которая оказывается постсинаптической по отношению к тормозному окончанию и пресинаптической по отношению к активируемой им нервной клетки. В окончаниях пресинаптического тормозного аксона освобождается медиатор, который вызывает деполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембраны для CI. Деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание аксона. В результате происходит угнетение процесса высвобождения медиатора возбуждающими нервными окончаниями и снижение амплитуды возбуждающего постсинаптического потенциала.
Характерной особенностью пресинаптической деполяризации является замедленное развитие и большая длительность (несколько сотен миллисекунд), даже после одиночного афферентного импульса.
Пресинаптическое торможение существенно отличается от постсинаптического и в фармакологическом отношении. Стрихнин и столбнячный токсин не влияют на его течение. Однако наркотизирующие вещества (хлоралоза, нембутал) значительно усиливают и удлиняют пресинаптическое торможение. Этот вид торможения обнаружен в различных отделах цнс. Наиболее часто оно выявляется в структурах мозгового ствола и спинного мозга. В первых исследованиях механизмов пресинаптического торможения считалось, что тормозное действие осуществляется в точке, отдаленной от сомы нейрона, поэтому его называли «отдаленным” торможением.
Функциональное значение пресинаптического торможения, охватывающего пресинаптические терминали, по которым поступают афферентные импульсы, заключается в ограничении поступления к нервным центрам афферентной импульсации. Пресинаптическое торможение в первую очередь блокирует слабые асинхронные афферентные сигналы и пропускает более сильные, следовательно, оно служит механизмом выделения, вычленения более интенсивных афферентных импульсов из общего потока. Это имеет огромное приспособительное значение для организма, так как из всех афферентных сигналов, идущих к нервным центрам, выделяются самые главные, самые необходимые для данного конкретного времени. Благодаря этому нервные центры, нервная система в целом освобождается от переработки менее существенной информации.
Вторичное торможение— торможение осуществляющееся теми же нервными структурами, в которых происходит возбуждение. Этот нервный процесс подробно изложен в работах Н.Е. Введенского (1886, 1901г.г.).
Торможение реципрокное (лат. reciprocus — взаимный) — нервный процесс, основанный на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через посредство вставочных нейронов торможение других групп клеток. Реципрокные отношения возбуждения и торможения в цнс были открыты и продемонстрированы Н.Е. Введенским: раздражение кожи на задней лапке у лягушки вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Взаимодействие возбуждения и торможения является общим свойством всей нервной системы и обнаруживается как в головном, так и в спинном мозге. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на взаимодействии возбуждения и торможения на одних и тех же нейронах цнс.
Общее центральное торможение —нервный процесс, развивающийся при любой рефлекторной деятельности и захватывавающий почти всю цнс, включая центры головного мозга. Общее центральное торможение обычно проявляется раньше возникновения какой-либо двигательной реакции. Оно может проявляться при такой малой силе раздражения при которой двигательный эффект отсутствует. Такого вида торможение было впервые описано И.С. Беритовым (1937). Оно обеспечивает концентрацию возбуждения других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражений. Важная роль в создании общего центрального торможения принадлежит желатинозной субстанции спинного мозга.
При электрическом раздражении желатинозной субстанции у спинального препарата кошки происходит общее торможение рефлекторных реакций, вызываемых раздражением сенсорных нервов. Общее торможение является важным фактором в создании целостной поведенческой деятельности животных, а также в обеспечении избирательного возбуждения определенных рабочих органов.
Парабиотическое торможениеразвивается при патологических состояниях, когда лабильность структур центральной нервной системы снижается или происходит очень массивное одновременное возбуждение большого числа афферентных путей, как, например, при травматическом шоке.
Некоторые исследователи выделяют еще один вид торможения — торможение вслед за возбуждением. Оно развивается в нейронах после окончания возбуждения в результате сильной следовой гиперполяризации мембраны (постсинаптической).
38. Внешнее торможение, его значение и виды.
Объяснить поведение, признавая лишь существование возбудительного процесса, нельзя. При наличии только процессов возбуждения возможны лишь судороги, а не целесообразная координированная деятельность. Функционирование условнорефлекторных механизмов основано на возбуждении и торможении. По мере упрочения условного рефлекса роль тормозного процесса возрастает. Торможение условнорефлекторной деятельности проявляется в форме внешнего, или безусловного, торможения и в форме внутреннего, или условного, торможения. Внешнее (безусловное) торможение — это врожденное генетически запрограммированное торможение. Различают два вида внешнего (безусловного) торможения: запредельное и индукционное.
Запредельное торможение условного рефлекса развивается либо при большой силе стимула, либо при слабом функционировании центральной нервной системы. Запредельное торможение имеет охранительное значение.
Индукционное (внешнее) торможение наблюдается в случае применения нового раздражителя после выработки условного рефлекса или наряду с известным условным раздражителем. На воздействие нового раздражителя будет осуществляться сильный врожденный ориентировочный рефлекс типа «Что такое?», направленный на оценку биологической значимости нового раздражителя.
39. Виды внутреннего торможения, их роль в процессе обучения.
Внутреннее торможение, в отличие от внешнего, развивается внутри дуги условного рефлекса, т. е. в тех нервных структурах, которые участвуют в осуществлении данного рефлекса.
Если внешнее торможение возникает сразу, как только подействовал тормозящий агент, то внутреннее торможение надо вырабатывать, оно возникает при определенных условиях, и это иногда требует длительного времени.
Одним из видов внутреннего торможения является угасание. Оно развивается, если много раз условный рефлекс не подкрепляется безусловным раздражителем.
Через некоторое время после угасания условный рефлекс может восстановиться. Это произойдет, если мы вновь подкрепим действие условного раздражителя безусловным.
Непрочные условные рефлексы восстанавливаются с трудом. Угасанием можно объяснить временную утрату трудового навыка, навыка игры на музыкальных инструментах.
У детей угасание происходит гораздо медленнее, чем у взрослых. Именно поэтому трудно отучать детей от вредных привычек. Угасание лежит в основе забывания.
Угасание условных рефлексов имеет важное биологическое значение. Благодаря ему организм перестает реагировать на сигналы, утратившие свое значение. Сколько бы ненужных, лишних движений при письме, трудовых операциях, спортивных упражнениях делал человек без угасательного торможения!
Запаздывание условных рефлексов также относится к внутреннему торможению. Оно развивается, если отставить во времени подкрепление условного раздражителя безусловным. ( Собака Павлова)
Запаздывание создает условия для лучшей ориентировки животного в окружающем мире.
Запаздывание у детей вырабатывается с большим трудом под влиянием воспитания и тренировки. Вспомните, как нетерпеливо тянет руку первоклассник, размахивая ею, вставая из-за парты, чтобы его заметил учитель. И только к старшему школьному возрасту мы отмечаем выдержку, умение сдерживать свои желания, силу воли.
Сходные звуковые, обонятельные и другие раздражители могут сигнализировать о совершенно различных событиях. Только точный анализ этих сходных раздражителей обеспечивает биологически целесообразные реакции животного. Анализ раздражений состоит в различении, разделении разных сигналов, дифференцировании сходных взаимодействий на организм.
Различение, или дифференцирование, сходных условных раздражителей вырабатывается путем подкрепления одних и неподкрепления других раздражителей. Развивающееся при этом торможение подавляет рефлекторную реакцию на неподкрепляемые раздражители. Дифференцировка — один из видов условного (внутреннего) торможения.
Благодаря дифференцировочному торможению можно выделить сигнально значимые признаки раздражителя из многих окружающих нас звуков, предметов, лиц и т. д. Дифференцирование вырабатывается у детей уже с первых месяцев жизни.
4. Безусловное торможение. Сущность внешнего и запредельного торможения. Условное торможение, его виды.
Торможение условного рефлекса:
Имеются 2 вида торможения условного рефлекса, принципиально отличающиеся др. от др.: врожденное и приобретеное, каждое из которых имеет собственные варианты.
Безусловное (врожденное) торможение:
1.внешнее;
2.запредельное.
Условное (приобретенное, внутреннее) торможение:
1.угасательное
2.запаздывательное
3.дифференцировочное
4.условный тормоз.
Внешнее торможение – появляется в ослаблении или прекращении наличного (протекающего в данный момент) условного рефлекса при действии какого-либо постороннего раздражителя.
Посторонний раздражитель à реакция, возникшая на изменение внешней среды (рефлекс на новизну) – рефлекс «что такое?»/ориентировочно исследовательская реакция (Павлов).
Эта реакция состоит из:
Настораживание – подготовка организма к действию на случай внезапно возникающей необходимости (нападение, бегство).
С повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, посколько организму не требуется предпринимать каких-то действий.
По степени выраженности влияния посторонних раздражителей на условно-рефлекторную деятельность выделяют 2 варианта торможения:
1. Гаснущий тормоз – посторонний сигнал, который с повторением его действия теряет свое тормозящее влияние, не имеет существенного значения для организма.
2. Постоянный тормоз – дополнительный раздражитель, с повторением не теряет своего тормозящего действия (пр: раздражения от переполненных внутренних органов, болевые).
Механизм внешнего торможения.: посторонний сигнал сопровождается появлением в КБМ нового очага возбуждения, который при средней силе раздражителя оказывает угнетающее влияние на текущую условно-рефлекторную деятельность по механизму доминанты.
Внешнее торможение — безусловно-рефлекторное.
Возбуждение клеток ориентировочно-исследовательского рефлекса, возникающее от постороннего раздражителя, находится вне дуги наличного условного рефлекса à поэтому – внешнее торможение.
Внешнее торможение способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды и дает возможность при необходимости переключится на другую деятельность в соответствии с ситуацией.
Запредельное торможение – возникает при длительном нервном возбуждении организма, при действии чрезвычайно сильного условного сигнала или нескольких несильных.
«закон силы»: чем сильнее условный сигнал, тем сильнее условно-рефлекторная реакция.
Закон силы сохраняется до определенной величины, выше которой эффект начинает уменьшаться, несмотря на увеличение силы условного сигнала: при достаточной силе условного сигнала эффект его действия может полностью исчезнуть => Корковые клетки имеют предел работоспособности.
Запредельное торможение по механизму часто относят к пессимильному, т.к. оно не требует специальной выработки, является безусловно-рефлекторным и выполняет охранительную роль.
Условное (приобретенное, внутренее) торможение – нервный процесс, требует своей выработки, как и сам рефлекс.
Является приобретенным и индивидуальным торможением = условно-рефлекторное торможение.
Локализовано в пределах нервного центра данного условного рефлекса.
Виды условного торможения:
Угасательное торможение – возникает при повторном применении условного сигнала и неподкреплены его.
Вначале условный рефлекс ослабевает, затем полностью исчезает, через некоторое время может восстановиться.
Чем значительнее интенсивность условного сигнала и биологическая значимость подкрепления, тем труднее совершается угасание условного рефлекса.
Если во время подкрепления условного угасательного рефлекса подействовать посторонним сигналом, возникает ориентировочно-исследовательский рефлекс, который ослабляет угасательное торможение и восстанавливает угасший ранее рефлекс (явление растормаживания).
Выработка угасательного торможения связана с активным угасанием условного рефлекса.
Угасший условный рефлекс быстро восстанавливается при его подкреплении.
Запаздывательное торможение – наступает тогда, когда подкрепление условного сигнала безусловным раздражителем осуществляется с большим опозданием (2—3 мин) по отношению к моменту предъявления условного раздражителя.
Дифференцировочное торможение – вырабатывается при дополнительном включении раздражителя, близкого к условному, и не подкреплении его.
Дифференцировочное торможение при действии посторонних сигналов средней силы ослабевает и сопровождается явлением растормаживания.
Условный тормоз – если условный стимул, на который образован условный рефлекс, применяется в комбинации с некоторым другим стимулом и их комбинация не подкрепляется безусловным стимулом, наступает торможение условного рефлекса, вызываемого этим стимулом.
Этот вид также растормаживается, если подключить другой раздражитель.
Значение всех видов условного торможения условных рефлексов–– в устранении ненужной в данное время деятельности – тонком приспособлении организма к окружающей среде.
При развитии условного торможения в этот процесс широко вовлекаются различные мозговые структуры.
Торможение условных рефлексов
Известно два вида торможения условных рефлексов, принципиально отличающихся друг от друга: врожденное (безусловное) и приобретенное (условное), каждое из которых имеет собственные варианты.
Торможение условных рефлексов

Безусловное
Внешнее
Запредельное
Условное (внутреннее)
Угасательное
Запаздывательное
Дифференцировочное
Условный тормоз
А. Врожденное (безусловное) торможение подразделяется на внешнее торможение и запредельное.
1. Внешнее торможение — это торможение, которое проявляется в ослаблении или прекращении наличного (протекающего в данный момент) условного рефлекса при действии какого-либо постороннего раздражителя. Например, включение звука, света во время текущего условного рефлекса вызывает появление ориентировочно-исследовательской реакции, которая ослабляет или прекращает наличную условнорефлекторную деятельность. Эту реакцию, возникшую на изменение окружающей среды (рефлекс на новизну), И. П. Павлов называл рефлексом «что такое?». Он состоит в настораживании и подготовке организма на случай внезапно возникающей необходимости к действию, например нападение, бегство. С повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, поскольку организму не требуется предпринимать каких-то действий.
По степени выраженности влияния посторонних раздражителей на условнорефлекторную деятельность выделяют два варианта торможения: гаснущий тормоз и постоянный тормоз. Гаснущий тормоз —это посторонний сигнал, который с повторением его действия теряет свое тормозящее влияние, поскольку не имеет существенного значения для организма. Обычно на человека действует масса различных сигналов, на которые сначала он обращает внимание, а затем перестает их «замечать». Постоянный тормоз — это такой дополнительный раздражитель, который с повторением не теряет своего тормозящего действия. Это раздражения от переполненных внутренних органов (например, от мочевого пузыря, кишечника), болевые раздражители. Они имеют существенное значение для человека и требуют от него принятия решительных мер к их устранению, поэтому условнорефлекторная деятельность затормаживается.
Механизм внешнего торможения. Согласно учению И.П.Павлова, посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, который при средней силе раздражителя оказывает угнетающее влияние на текущую условнорефлекторную деятельность по механизму доминанты. Внешнее торможение является безусловнорефлекторным. Поскольку в этих случаях возбуждение клеток ориентировочно-исследовательского рефлекса, возникающего от постороннего раздражителя, находится вне дуги наличного условного рефлекса, это торможение назвали внешним. Более сильный или более важный в биологическом или социальном отношении раздражитель подавляет (ослабляет или устраняет) другую реакцию. Внешнее торможение способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды организма и дает возможность при необходимости переключиться на другую деятельность согласно ситуации.
2. Запредельное торможение возникает при действии чрезвычайно сильного условного сигнала. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие — «закон силы»: чем сильнее условный сигнал, тем сильнее условнорефлекторная реакция. Однако закон силы сохраняется до определенной величины, выше которой эффект начинает уменьшаться, несмотря на увеличение силы условного сигнала: при достаточной силе условного сигнала эффект его действия может полностью исчезнуть. Эти факты позволили И.П.Павлову выдвинуть представление о том, что корковые клетки имеют предел работоспособности. Многие исследователи относят запредельное торможение по механизму к пессимальному торможению (угнетение деятельности нейрона при чрезмерно частом его возбуждении, превышающем лабильность). Поскольку появление указанного торможения не требует специальной выработки, оно, как и внешнее торможение, является безусловнорефлекторным.
Б. Условное торможение условных рефлексов (приобретенное, внутреннее) требует своей выработки, как и сам рефлекс. Поэтому его и называют условнорефлекторным торможением: оно является приобретенным, индивидуальным. Согласно учению И.П.Павлова, оно локализовано в пределах («внутри») нервного центра данного условного рефлекса. Различают следующие виды условного торможения: угасательное, запаздывательное, дифференцировочное и условный тормоз.
11. Угасателъное торможение возникает при повторном применении условного сигнала и неподкреплении его. При этом вначале условный рефлекс ослабевает, а затем полностью исчезает. Через некоторое время он может восстановиться. Скорость угасания зависит от интенсивности условного сигнала и биологической значимости подкрепления: чем они значительнее, тем труднее совершается угасание условного рефлекса. Этот процесс связан с забыванием полученной ранее информации, если она длительно не повторяется. Если во время проявления условного угасательного рефлекса действует посторонний сигнал, возникает ориентировочно-исследовательский рефлекс, который ослабляет угасательное торможение и восстанавливает угасший ранее рефлекс (явление растормаживания). Это показывает, что выработка угасательного торможения связана с активным угасанием условного рефлекса. Угасший условный рефлекс быстро восстанавливается при его подкреплении.
Запаздывателъное торможение возникает при отставлении подкрепления на 1-3 мин относительно начала действия условного сигнала. Постепенно появление условной реакции сдвигается к моменту подкрепления. Более длительное отставление подкрепления в опытах на собаках не удается. Выработка запаздывательного условного торможения наиболее трудна. Этому торможению также присуще явление растормаживания.
Дифференцировочное торможение вырабатывается при дополнительном включении раздражителя, близкого к условному, и неподкреплении его. Например, если у собаки тон 500 Гц подкреплять пищей, а тон 1000 Гц не подкреплять и чередовать их в течение каждого опыта, то через некоторое время животное начинает различать оба сигнала: на тон 500 Гц возникнет условный рефлекс в виде движения к кормушке, поедания корма, слюноотделения, а на тон 1000 Гц животное будет отворачиваться от кормушки с пищей, слюноотделение не появится. Чем меньше различия между сигналами, тем труднее идет выработка дифференцировочного торможения. У животных удается выработать различение частот метронома — 100 и 104 удара/мин, тонов 1000 и 995 Гц, распознавание геометрических фигур, различение раздражения разных участков кожи и т.д. Условное дифференцировочное торможение при действии посторонних сигналов средней силы ослабевает и сопровождается явлением растормаживания, т.е. это такой же активный процесс, как и придругих видах условного торможения.
Условный тормоз возникает при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Если выработать, например, условный слюноотделительный рефлекс на свет и затем к условному сигналу «свет» подключить дополнительный раздражитель, например «звонок», не подкреплять эту комбинацию, то постепенно условный рефлекс на нее угасает. Сигнал «свет» необходимо продолжать подкреплять пищей или вливанием в рот слабого раствора кислоты. После этого присоединение сигнала «звонок» к любому условному рефлексу ослабляет его, т.е. «звонок» стал условным тормозом для любого условного рефлекса. Этот вид торможения также растормаживается, если подключить другой раздражитель.
Функциональные изменения при выработке условных рефлексов и условного торможения (изменения возбудимости, ЦНС, ЭЭГ) имеют общие черты, так же как одинаковы стадии их формирования. Условное торможение называют также отрицательным условным рефлексом.
Значение всех видов условного (внутреннего) торможения условных рефлексов заключается в устранении ненужной в данное время деятельности — тонком приспособлении организма к окружающей среде.
22. Внешнее и внутреннее торможение условных рефлексов по и.П. Павлову. Виды внутреннего торможения. Запредельное торможение.
Условные рефлексы не только вырабатываются, но и исчезают при определенных условиях. И. П. Павлов различал два вида торможения условных рефлексов: безусловное и условное.
Безусловное торможение является врожденным, оно может проявляться в любом отделе ЦНС. Безусловное торможение может быть внешним и запредельным. Внешнее торможение возникает под влиянием нового раздражителя, который действует одновременно с условным сигналом. Внешний раздражитель должен быть более сильным — доминантным. Например, болевое раздражение кожи у собаки может резко затормозить пищевые условные рефлексы. Положительное значение внешнего торможения состоит в том, что организм переключается на новый, более важный в данный момент, вид рефлекторной деятельности.
Запредельное торможение возникает при значительном увеличении силы или продолжительности действия условного сигнала. При этом условный рефлекс резко ослабевает или полностью исчезает. Например, у собаки был выработан слюноотделительный условный рефлекс на звонок. Если постепенно увеличивать силу условного сигнала (звонка), то вначале количество отделяемой слюны увеличивается. При дальнейшем нарастании силы условного сигнала отделение слюны уменьшается и, наконец, полностью тормозится.
По своей природе запредельное торможение является пессимальным. Оно выполняет охранительную функцию, препятствуя истощению нервных клеток.Запредельное торможение легче развивается при снижении лабильности, работоспособности нейронов коры большого мозга, например, после тяжелого инфекционного заболевания, у пожилых людей и т. д.
Условное (внутреннее) торможение свойственно только клеткам КГМ. Это торможение, как и условные рефлексы, вырабатывается. Основным условием для проявления внутреннего торможения является неподкрепление условного раздражителя безусловным. Например, если у собаки выработан прочный слюноотделительный условный рефлекс на свет, а затем условный сигнал (свет) применять много раз изолированно без подкрепления (без дачи пищи), то выделение слюны постепенно уменьшается и, наконец, прекратится. Условный рефлекс угас — угасательное торможение. Подкрепление условного сигнала безусловным раздражителем восстанавливает условный рефлекс. Однако даже при отсутствии подкрепления условный рефлекс может вновь проявиться после отдыха, при наличии положительных эмоций. Это явление получило название растормаживания условных рефлексов. Быстрее и легче угасают непрочные недавно выработанные условные рефлексы. За счет угасательного торможения организм освобождается от ненужных, потерявших сигнальное значение условных рефлексов.
Значение торможения условных рефлексов. За счет торможения условных рефлексов достигается точное и совершенное приспособление организма к условиям существования, уравновешивание организма с окружающей средой, осуществляется аналитическая и синтетическая деятельность головного мозга.
В клетках высших отделов, мозга развивается охранительное, или запредельное, торможение, возникающее под влиянием чрезмерно длительных, очень сильных или частых раздражений. Такие раздражители как бы превышают предел возможной работоспособности нервных клеток. Охранительное торможение предохраняет клетки от функционального разрушения. В нейронах коры больших полушарий мозга, имеющих низкую функциональную подвижность, охранительное торможение развивается относительно легко. Охранительное торможение может быть вызвано действием частых проприорецептивных импульсов от работающих мышц при беге, на короткие дистанции с максимальной скоростью. Примером запредельного торможения служит стартовая апатия ( XIX), возникающая у спортсменов перед стартом и отрицательно влияющая на спортивный результат. Интенсивность и скорость развития охранительного торможения, зависят от возраста, индивидуальных особенностей, состояния нервных клеток и условий предшествующей деятельности. Условное (внутреннее) торможение, в отличие от безусловного, вырабатывается при определенных условиях, постепенно. Оно временно и является активным процессом. Этот вид торможения свойствен только высшим отделам головного мозга. В зависимости от условий возникновения различают: угасательное, дифференцировочное и запаздывающее торможение. Угасательное торможение возникает и развивается вследствие неподкрепления условного раздражителя (сигнала) безусловным, или прочным условным рефлексом. Прочно выработанные условные рефлексы и рефлексы, полученные при сильном подкреплении, угашаются медленно. Быстрота угасания зависит от частоты, неподкрепления, т. е. чем чаще не подкрепляется условный раздражитель, тем быстрее угасает рефлекс. Чем глубже угасание, тем большее влияние оказывает оно на величину других условных рефлексов и даже на безусловных.