Виды торможения: 403 — Доступ запрещён – Способы торможения автомобиля
- 09.07.2020
9. Первичное торможение. Его виды. Механизм возникновения
Характеристика первичного торможения В центральной нервной системе тормозные нейроны есть в спинном мозге, в головном мозге (меньшее количество) и в коре головного мозга (большинство). В спинном мозге 2 вида тормозных нейронов (это тормозные вставочные нейроны):
клетки Реншоу — не обладают фоновой активностью и в покое не генерируют нервных импульсов. Они возбуждаются под действием: импульсов от афферентных нейронов, от эфферентных нейронов (альфа-мотонейронов спинного мозга), импульсов от вышележащих отделов головного мозга;
клетки Уилсона — обладают постоянной фоновой активностью, даже без раздражения (в покое) они генерируют нервные импульсы — постоянно тормозят активность альфа-мотонейронов спинного мозга.
В коре головного мозга 4 вида тормозных клеток:
большие корзинчатые нейроны — 3, 4, 5 слои коры головного мозга, их аксоны сильно ветвятся и образуют сплетения на площади около 500 мкм. Они тормозят активность нейронов 3, 4, 5 слоев;
малые корзинчатые клетки нейроны — 2, 3 слои коры — их аксоны ветвятся на меньшей площади, около 50 мкм, и тормозят 2 и 3 слои;
нейроны с кистеобразным аксоном — 1 слой коры, образует аксон, на конце которого разветвление в виде кисти; тормозит клетки 1-го слоя;
канделяброобразные нейроны — на границе между 2 и 3 слоями, вниз от них идет аксон и дает несколько ответвлений вверх, тормозят активность всех слоев.
Первичное торможение осуществляется за счет выделения тормозного медиатора на окончаниях нервных клеток (ГАМК — гаммааминомаслянная кислота, глицин, таурин, серотонин и другие). Эти вещества вызывают гиперполяризацию постсинаптической мембраны и, как следствие, тормозной постсинаптический потенциал. Различают 2 вида первичного торможения.
Пресинаптическое — развивается на мембране возбужденного синапса. Развивается в аксо-аксональном синапсе. Медиатор — гаммааминомаслянная кислота — он изменяет проницаемость клеточной мембраны для CL- и Са2+ (понижает ее). В результате на постсинаптической мембране тормозного синапса возникает явление стойкой деполяризации, затем — падение возбудимости и возбуждение по аксону не доходит до альфа-мотонейрона — блок проведения возбуждения. За счет снижение проницаемости для Са2+ снижается количество медиатора в возбужденном синапсе и, как следствие, на теле иннервируемой клетки нет возбуждающего постсинаптического потенциала.
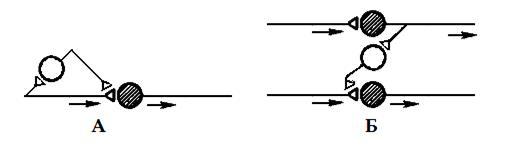
Рис. 4. Разновидности пресинаптического торможения: А – параллельное, Б – латеральное. Темные нейроны – возбуждающие, светлые – тормозные. Постсинаптическое торможение — обеспечивается за счет гаммааминомасляной кислоты и глицина. Тормозная клетка образует синапс на теле нейрона. На окончании тормозного нейрона выделяется тормозной медиатор, который вызывает гиперполяризацию постсинаптической мембраны. Возникает тормозной постсинаптический потенциал и величина возникшего постсинаптического потенциала уменьшается. Постсинаптическое торможение активно используется в нейронных сетях, и в зависимости от вариантов связывания нейронов друг с другом выделяют несколько его видов:
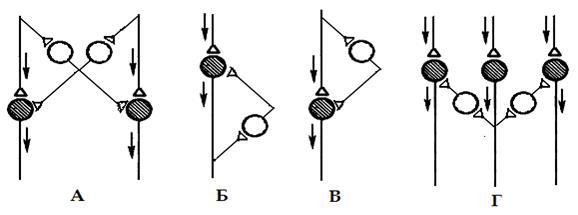
Рис. 3. Разновидности постсинаптического торможения: А – реципрокное, Б – возвратное, В – параллельное, Г- латеральное. Темные нейроны – возбуждающие, светлые – тормозные
Реципрокное торможение
Возвратное торможение (рис. 3.Б) — это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам. Возвратное торможение наблюдается, например, в мотонейронах спинного мозга позвоночных. Эти клетки отдают возвратные коллатерали в мозг к тормозным вставочным клеткам Реншоу, которые имеют синапсы на этих же мотонейронах. Торможение обеспечивает ограничение ритма мотонейронов, позволяющее чередовать сокращение и расслабление скелетной мышцы, что важно для нормальной работы двигательного аппарата.
Такую же роль играет возвратное торможение и в других нервных сетях.
Параллельное торможение (рис. 3.В) – играет сходную с возвратным роль, но в этом случае возбуждение блокирует само себя, посылая тормозной сигнал на нейрон который одновременно и активирует.
Это возможно, если возбуждающий импульс сам не должен вызвать возбуждения на нейроне-мишени, но его роль важна при пространственной суммации, в комбинации с другими сигналами.
Латеральное торможение (рис. 3.Г) – это торможение нервных клеток, расположенных по соседству с активной, которое этой клеткой и инициируется. При этом вокруг возбужденного нейрона возникает зона, в которой развивается очень глубокое торможение.
Латеральное торможение наблюдается, например, в конкурирующих сенсорных каналах связи. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. Во всех случаях латеральное торможение обеспечивает
Принципы координационной деятельности ЦНС (конвергенция, общий конечный пункт, дивергенция, иррадиация, реципрокность, доминанта).
Нервная система человека и животных может быть представлена как система нейронных цепочек, передающих возбуждающие и тормозные сигналы (нервная сеть). Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путем регулировки их входов. Такие нейронные цепи построены из стандартных элементов, которые выполняют наиболее часто повторяющиеся операции и могут быть включены в схемы самых разнообразных нервных структур.
Существуют значительные количественные различия нервных сетей у разных видов позвоночных и беспозвоночных. Так, у человека нервная система включает около 1010элементов, у примитивных беспозвоночных — около 104 нейронов, Однако в строении и функционировании всех нервных систем имеются общие черты. Практически во всех отделах центральной нервной системы обнаружены дивергенция нервных путей, конвергенция нервных путей и различные варианты тормозных связей между элементами нервных цепочек.
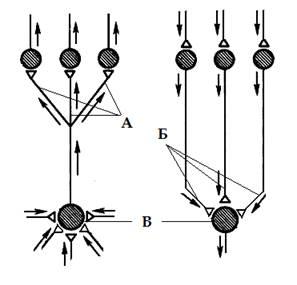
Рис 2. Дивергенция (А), конвергенция (Б) и пространственная суммация (В) нервных путей в центральной нервной системе.
Дивергенция и конвергенция путей.
Дивергенция (расхождение) пути (рис 2.А) — возникает в результате контактирования одного нейрона с множеством нейронов более высоких порядков. Так, например, происходит разделение аксона чувствительного нейрона, входящего в спинной мозг, на множество веточек (коллатералей), которые направляются к разным сегментам спинного мозга и в головной мозг, где происходит передача сигнала на вставочные и далее — на моторные нервные клетки. Дивергенция пути сигнала наблюдается так же у вставочных и у эффекторных нейронов.
Дивергенция пути обеспечивает расширение сферы действия сигнала, благодаря ей, информация поступает одновременно к разным участкам ЦНС. Это называют иррадиациейвозбуждения (или торможения). Дивергенция настолько обычное явление, что можно говорить о принципе дивергенции в нейронных цепях.
Конвергенция — это схождение многих нервных путей к одним и тем же нейронам (рис 2.Б). Например, у позвоночных на каждом мотонейроне спинного мозга и ствола головного мозга образуют нервные окончания тысячи сенсорных, а также возбуждающих и тормозных вставочных нейронов разных уровней. Мощная конвергенция обнаруживается и на нейронах ретикулярной формации ствола мозга, на многих корковых нейронах у позвоночных и, видимо, на командных нейронах.
Конвергенция многих нервных путей к одному нейрону делает этот нейрон интегратором соответствующих сигналов. Вероятность возбуждения такого нейрона-интегратора зависит не от каждого пришедшего стимула в отдельности, а от суммы и направления стимулов, действующих одновременно, то есть суммы всех синаптических процессов происходящих на его плазматической мембране. Другими словами, вероятность распространения возбуждения через нейрон-интегратор определяется алгебраическим сложением величин возбуждающих и тормозных входов на нем, активных в данный момент. Такое сложение является результатом или пространственной или временной суммации.
Пространственная суммация – результат сложения нервных импульсов приходящих одновременно к нейрону через разные синапсы (рис 2.В), временная суммация – сложение приходящих поочередно, через один синапс с небольшими интервалами времени. В обоих случаях нейрон интегратор, называют общим путем для конвергирующих на него нервных сигналов, а если речь идет о мотонейроне, т. е. конечном звене нервного пути к мускулатуре, говорят об общем конечном пути.
Результат суммации заключается в возможности изменения направления распространения возбуждения в ЦНС, (то есть не строго в пределах одной рефлекторной дуги), а значит и в изменении характера ответной реакции организма в ответ на действие раздражителя. Ответ организма, реализуемый в результате, становится более адекватным внешним условиям и состоянию нервной системы. Пример такого выбора ответа можно видеть, если речь идет о конвергенции не на одном нейроне, а на группе нейронов совместно регулирующих общую функцию, что в ЦНС не редкость. Наличие конвергенции множества путей на одной группе мотонейронов лежит в основе феноменов пространственного облегчения и окклюзии.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.
Принцип реципрокности (сопряженности) в работе нервных центров. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов — сгибателей и разгибателей конечностей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы.
Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное), но может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр.
Принцип общего конечного пути
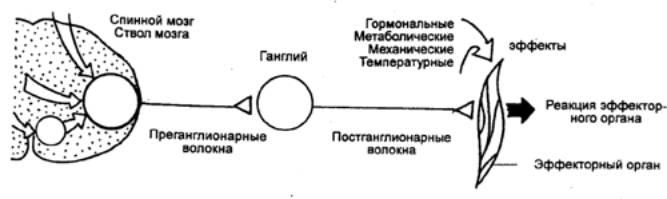
В структурной организации нервных сетей встречается такая ситуация, когда на одном нейроне сходятся несколько афферентных терминалей из других отделов ЦНС (рис. 3.2). Это явление принято называтьконвергенцией в нейронных связях. Так, например, к одному мотонейрону подходит около 6000 коллатералей аксонов первичных афферентов, спинальных интернейронов, нисходящих путей из стволовой части мозга и коры. Все эти терминальные окончания образуют на мотонейроне возбуждающие и тормозные синапсы и формируют своеобразную «воронку», суженная часть которой представляет общий моторный выход.Данная воронка является анатомическим образованием, определяющим один из механизмов координационной функции спинного мозга (рис. 3.3).
Суть этого механизма была раскрыта английским физиологом Ч. Шеррингтоном, который сформулировал принцип общего конечного пути. По Ч. Шеррингтону, количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути, которым являются группа мотонейронов и иннервируемые ими мышцы. В результате этого столкновения достигается торможение всех возможных степеней свободы двигательного аппарата, кроме одной, в направлении которой протекает рефлекторная реакция, вызванная максимальной стимуляцией одного из афферентных входов.
Рассмотрим случай с одновременным раздражением рецептивных полей чесательного и сгибательного рефлексов, которые реализуются идентичными группами мышц. Импульсы, идущие от этих рецептивных полей, приходят к одной и той же группе мотонейронов, и здесь в узком месте воронки за счет интеграции синаптических влияний осуществляется выбор в пользу сгибательного рефлекса, вызванного более сильным болевым раздражением. Принцип общего конечного пути, как один из принципов координации, действителен не только для спинного мозга, он применим для любого этажа ЦНС, в том числе для моторной коры.
|
Рис. 3.3 Нейронный субстрат общего конечного пути автономной нервной системы |
Принцип доминанты. Принцип доминанты сформулирован А. Л. Ухтомским, Этот принцип играет важную роль в согласованной работе нервных центров. Доминанта — это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами:
· повышенной возбудимостью;
· стойкостью возбуждения;
· способностью к суммированию возбуждения;
· инерцией, доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
124. Явление торможения в высшей нервной деятельности. Виды торможения. Современное представление о механизмах торможения.
Изучая закономерности ВНД, И.П. Павлов установил, что существует 2 вида торможения условных рефлексов: внешнее (безусловное) и внутреннее (условное). Внешнее торможение – это процесс экстренного ослабления или прекращения условнорефлекторных реакций в результате действия посторонних раздражителей, т.е. стимулов или сигналов, поступающих из внешней или внутренней среды. Безусловным это раздражение называется потому, что является врождённым и не требует выработки.
Существует 2 разновидности безусловного торможения:
1.Внешний тормоз. Он делится в свою очередь на постоянный тормоз и гаснущий тормоз. Постоянным тормозом называются такие посторонние стимулы, которые всегда вызывают торможение условных рефлексов. Например, болевые раздражители. Гаснущим тормозом называются стимулы, тормозящий эффект которых с течением времени начинает уменьшаться. Например, если во время условнорефлекторного слюноотделения на звонок, появляется другой звук, то у собаки возникает ориентировочный рефлекс, поэтому слюноотделение тормозится. Однако спустя некоторое время ориентировочная реакция исчезает, а слюноотделение продолжается.
2.Запредельное торможение. Оно развивается при действии очень сильных раздражителей или длительном воздействии умеренных стимулов. В этом случае возникающее возбуждение нейронов коры превышает предел их работоспособности. В результате запредельного торможения нейроны временно выключаются для восстановления возбудимости и работоспособности. Поэтому данный вид торможения ещё называют охранительным. Например у некоторых людей сильные эмоции вызывают состояние заторможенности. Этот вид торможения является одной из основ закона силовых отношений. Согласно этому закону, чем сильнее раздражитель, тем более выражена рефлекторная реакция. Однако при сверхсильных стимулах из-за запредельного торможения она прекращается.
Таким образом, безусловное торможение выполняет две основные функции:
1.Координационную, т.е. способствует выключению биологически несущественных для данной ситуации процессов возбуждения в коре.
2.Охранительную, предупреждая истощение и гибель клеток коры.
Условное торможение — это торможение условных рефлексов, возникающее в результате выработки. Его называют также внутренним. Существуют следующие виды условного торможения:
1.Угасательное торможение. Оно возникает в том случае, если после выработки устойчивого условного рефлекса прекратить дальнейшее подкрепление условного раздражителя безусловным. Через некоторое время реакция на условный раздражитель исчезнет. Значение угасательного торможения заключается в выключении условных рефлексов, потерявших своё значение. Условнорефлекторная связь не разрывается, а только тормозится. Поэтому если во время действия условнорефлекторного стимула, на который реакция исчезла, подействовать сильным посторонним раздражителем, то может произойти растормаживание угашенного рефлекса. Это используется в клинике, для восстановления памяти, речи.
2.Дифференцированное торможение. Оно возникает, когда на сенсорные системы действует группа близких по характеру условных раздражителя. Например звуки близкие по частоте. При этом один из них подкрепляется безусловнорефлекторным стимулом, а остальные нет. Первоначально условнорефлекторная реакция будет возникать на все похожие раздражения, а спустя некоторое время только на тот, который подкрепляется. Дифференцировка, т.е. различение раздражителей развивается тем быстрее, чем меньше сходство между ними и наоборот. Дифференцировочное торможение обеспечивает выделение нужных сигналов. Наиболее высока способность к дифференцировке у человека. Однако она ухудшается при наличии сильных или множественных внешних сигналов. Например, умственная работа в условиях шума.
3.Запаздывательное торможение. Оно наблюдается в том случае, если постепенно время действия безусловного подкрепляющего раздражителя отодвигать от момента включения условного. Постепенно условнорефлекторная реакция также сдвигается к времени действия подкрепляющего раздражителя. С помощью этого вида торможения рефлекторная реакция сдвигается ближе к моменту подкрепления. Это способствует экономной работе нейронов мозга. У человека многие рефлекса являются запаздывающими, причём у возбудимых людей запаздывательное торможение вырабатывается труднее (необдуманные поступки).
4.Условный тормоз. Он возникает тогда, когда условнорефлекторный раздражитель подкрепляется, а его комбинации с другим раздражителем нет. Первоначально реакция возникает и на комбинацию двух раздражителей. Но затем только на один условнорефлекторный. Значение условного тормоза, как и дифференцировки состоит в различении сигналов.
Активация механизмов внутреннего торможения используется для гипноза. При монотонном многократном воздействии слабых раздражителей развивается внутреннее торможение (метроном, блестящий шарик, монотонная речь).
17. Виды торможения, взаимодействие процессов возбуждения и торможения в ЦНС
17. Виды торможения, взаимодействие процессов возбуждения и торможения в ЦНС
Торможение – активный процесс, возникающий при действии раздражителей на ткань, проявляется в подавлении другого возбуждения, функционального отправления ткани нет.
Торможение может развиваться только в форме локального ответа.
Выделяют два типа торможения:
1) первичное. Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения под воздействием тормозного медиатора. Различают два вида первичного торможения:
а) пресинаптическое в аксо-аксональном синапсе;
б) постсинаптическое в аксодендрическом синапсе.
2) вторичное. Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. Виды вторичного торможения:
а) запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона;
б) пессимальное, возникающее при высокой частоте раздражения;
в) парабиотическое, возникающее при сильно и длительно действующем раздражении;
г) торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения;
д) торможение по принципу отрицательной индукции;
е) торможение условных рефлексов.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса.
Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Торможение в ЦНС может возникать при одновременном поступлении в спинной мозг нервных импульсов различной силы с нескольких раздражителей. Более сильное раздражение тормозит рефлексы, которые должны были наступать в ответ на более слабые.
В 1862 г. И. М. Сеченов доказал в своем опыте, что раздражение кристалликом хлорида натрия зрительных бугров лягушки вызывает торможение рефлексов спинного мозга. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась.
Результат этого опыта позволил И. М. Сеченому сделать заключение, что в ЦНС наряду с процессом возбуждения развивается процесс торможения, который способен угнетать рефлекторные акты организма.
Поделитесь на страничкеСледующая глава >
4.8. Процесс торможения в цнс
Торможение – это активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение вторично относительно процесса возбуждения, так как всегда возникает как его следствие.
Торможение в ЦНС открыл И.М. Сеченов в 1863 г. В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Было показано, что латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Открытие И.М. Сеченова послужило толчком для дальнейших исследований торможения в ЦНС.
По месту возникновения различают пост- и пресинаптическое раздражение.
4.8.1. Постсинаптическое торможение
Постсинаптическое торможение открыл Дж. Экклс (1952) при Регистрации потенциалов мотонейронов спинного мозга у кошки во время раздражения мышечных афферентов. При этом оказалось, Что в мотонейронах мышцы-антогониста регистрируется не деполяризация, а гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона, угнетающий его способность реагировать на возбуждающие влияния. По этой причине вызванный гиперполяризационный потенциал был назван тормозным постсинаптическим потенциалом – ТПСП (рис. 4.9).
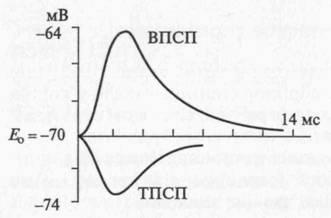
Рис. 4.9. Возбуждающий (ВПСП) и тормозной (ТПСП) постсинаптические потенциалы
У кошки ТПСП регистрируется на 0,5 мс позже, чем ВПСП, что объясняется наличием на пути проведения возбуждения, запускающего ТПСП, одного дополнительного синапса. Амплитуда ТПСП 1–5 мВ, он способен суммироваться, более мощный афферентный залп вызывает возрастание амплитуды ТПСП.
Механизм постсинаптического торможения. ТПСП уменьшает возбудимость клетки, т.е. увеличивает пороговый потенциал (AV), так как Екр (критический уровень деполяризации – КУД) остается на прежнем уровне, а мембранный потенциал (Ео) возрастает. ТПСП возникает под влиянием аминокислоты глицина, а также гамма-аминомасляной кислоты (ГАМК). В спинном мозге глицин выделяется особыми тормозными клетками (клетками Реншоу) в синапсах, образуемых этими клетками на мембране нейрона-мишени. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для Сl–, при этом Сl– поступает в клетку согласно концентрационному градиенту и вопреки электрическому градиенту, в результате чего развивается гиперполяризация. В среде, обедненной хлором, тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем в зоне аксонного холмика не происходит выведения, мембранный потенциал не достигает критического уровня. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа Сl– в клетку или выхода К+ из клетки.
Концентрационные градиенты ионов К+ в процессе развития торможения нейронов поддерживаются Na+/К+-помпой, ионов Сl––Сl–-помпой
Разновидности постсинаптического торможения. Обычно выделяют возвратное, латеральное, параллельное и прямое (реципрокное) постсинаптическое торможение. Имеются и другие варианты классификаций. Некоторые авторы называют только два вида торможения: возвратное и прямое (трактуется по-разному). В реальности вариантов торможения больше: они определяются множеством связей различных нейронов, в частности их коллатералей.
Возвратное постсинаптическое торможение – это такое торможение, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. В этом случае развивающееся торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Типичным примером возвратного постсинаптического торможения является торможение в мотонейронах спинного мозга. Как следует из рис. 4.10, 2, мотонейроны посылают коллатерали к тормозным вставочным нейронам, аксоны которых, в свою очередь, образуют синапсы на тех же мотонейронах, которые возбуждают тормозную клетку Реншоу. Такая тормозная цепь называется торможением Реншоу (в честь ученого, который ее открыл), а тормозные вставочные нейроны в этой цепи – клетками Реншоу. Это торможение обеспечивает, например, поочередное сокращение и расслабление скелетных мышц – сгибателей и разгибателей, что необходимо для координации движений конечностей при ходьбе. Сама клетка Реншоу возбуждается под влиянием ацетилхолина, воздействующего на Н-холинорецептор.
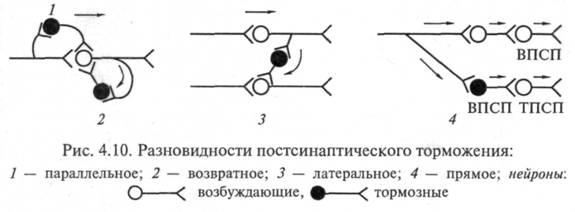
Подобную роль может выполнять и параллельное торможение, когда возбуждение блокирует само себя за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением (рис 4.10, 7).
Латеральное постсинаптическое торможение графически представлено на рис. 4.11, 3. Тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в этих соседних клетках развивается очень глубокое торможение, называемое латеральным, так как образующаяся зона торможения находится «сбоку» по отношению к возбужденному нейрону и инициируется им. Латеральное торможение играет особенно важную роль в афферентных системах: оно может образовать тормозную зону, которая окружает возбуждающие нейроны.

Примером прямого торможения может служить реципрокное торможение, вызывающее угнетение центра-антагониста. Например, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс: центр сгибания возбужден, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку Реншоу – к центру мышцы-антагониста, т.е. разгибателю, что предотвращает ее сокращение (рис. 4.10, 4). Если бы возбуждались одновременно центры мышц-сгибателей и мышц-разгибателей, сгибание конечности в суставе было бы невозможным.